Bestseller
Hello US customer, you are currently shopping on the UK site. To ensure you are charged in the correct currency and receive accurate shipping costs, please switch to the US website and re-add your product to your basket.

The UK's No. 1 bestselling brand
of friendly bacteria supplements^
.png?lang=en-GB)
Rated 'Excellent' on Trustpilot
4.8 stars (based on 5,721 reviews)

Scientifically proven to reach the gut
& complement the natural microbiome
FREE UK DELIVERY on orders over £40
and 14-day hassle-free returns
You're away from FREE UK delivery Free UK delivery (applied at checkout) on orders over £40.00
You have qualified for Free UK delivery

At Optibac we specialise entirely in live cultures and have many different supplements for different life stages and individual needs.
So you can feel good about choosing Optibac.
We only use the most highly researched strains of friendly bacteria

We prioritise natural ingredients and take care to avoid hidden sugars and other nasties

From over 5,000 independently verified reviews on Trustpilot

We have specialised in creating friendly bacteria supplements for over 20 years.

Our specialist range is designed to target individual needs and life stages

Including glass jars, compostable pouches and FSC®-certified cartons

"When you consider the quality of the strains used, and the billions per capsule, Optibac actually offers incredible value for money!"

"I love that the range is tailored for different life stages and needs. It takes into account our individuality - which is important."

"Most people don’t know that the research behind the strains is so much more important than a higher billion count per capsule."

"Optibac use more natural ingredients and fewer additives than most brands. The gummies are made from fruit purees with no added sugar or flavourings – that’s rare!"

"The Optibac range is home to the most researched strains in the world. It feels good being able to recommend supplements that you genuinely believe in!"

"My personal discovery of live cultures started with ‘One Week Flat’ 10 years ago and it’s one I still use and recommend regularly, especially before holidays or festive periods."
We specialise in friendly bacteria and have created a unique range of supplements designed to meet the needs of the whole family. Our supplements are proven to not only reach the gut alive, but to complement the natural microbiome. We genuinely care about helping the planet and its people, and so far have raised £1.5 million for charitable organisations such as Womankind Worldwide.
More about us
We are truly honoured to have been awarded the Queen's Award for Enterprise. This is one of the most prestigious awards for business in the UK, designed to celebrate the best of British business and recognise companies who are making an outstanding contribution to enterprise. In 2019 we won in the International Trade category.

We care about helping our customers make the best possible choice from the friendly bacteria supplements in our range. Sometimes you need a little help, so we've made it as easy as possible to settle on the right friendly bacteria for your needs.
Get StartedAt Optibac we specialise entirely in good bacteria and we understand the importance of using specific strains for your individual needs.
Learn about specific strainsThis is the most researched strain of L. acidophilus in the world and has undergone numerous large scale clinical trials. Find it in our high-strength supplements, Every Day EXTRA and Every Day MAX.
This strain of friendly yeast has been used safely around the world for over 50 years. It’s very robust and is resistant to heat, humidity and stomach acidity. Find it in our S Boulardii.
This is the most researched strain of friendly bacteria in the world. It has been extensively studied around the world in all age groups, and we're proud to offer it in our Babies & Children supplement.
One of the most researched strains of Bacillus, trialled in hundreds of children and adults. You can find it in our gut health gummy supplements, Gut Health Gummies and Kids Gummies.
This highly studied strain for the intimate flora has been trialled in thousands of women around the world. It can be found in For Women, the UK's No 1 friendly bacteria supplement for the vagina^^.
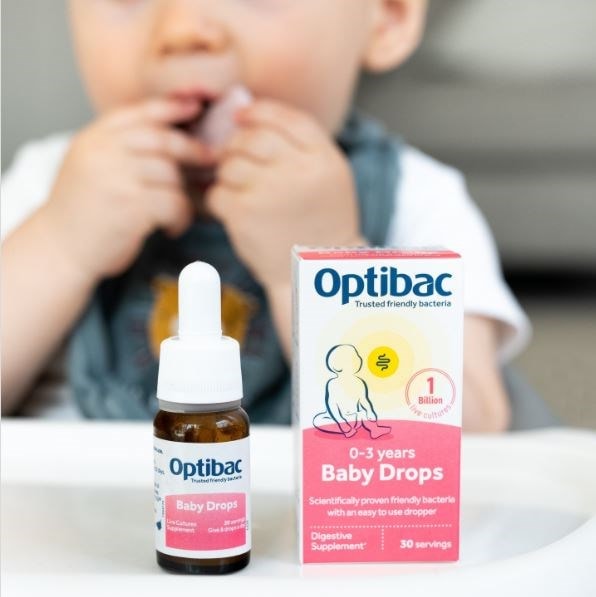
This safe, highly researched strain has been clinically trialled in over 4,000 infants and young children, including newborns. You can find it in our Baby Drops and Babies & Children.
| Bio-Kult | Symprove | ||
|---|---|---|---|
| No. of clinical trials | No. of clinical trials 26 | No. of clinical trials 19 | No. of clinical trials 3 |
| Eco conscious packaging | Eco conscious packaging Yes | Eco conscious packaging No | Eco conscious packaging No |
| Daily recommendation | Daily recommendation 1 capsule | Daily recommendation 1 - 4 capsules | Daily recommendation 70ml shot |
| Price | Price £12.99 | Price £10.48 | Price £79.00 |
| Price per day | Price per day 43p | Price per day 35p to £1.40 | Price per day £2.63 |
Comparing Optibac Every Day 30 capsules, Bio-Kult Everyday (formerly Advanced Multi Strain Formulation) 30 capsules and Symprove 500ml bottle in either flavour. Clinical trials counted across strains or combination of strains in each product. Optibac Every Day packaging is widely recyclable and not in plastic containers nor blister packs.
Click to browse the full range